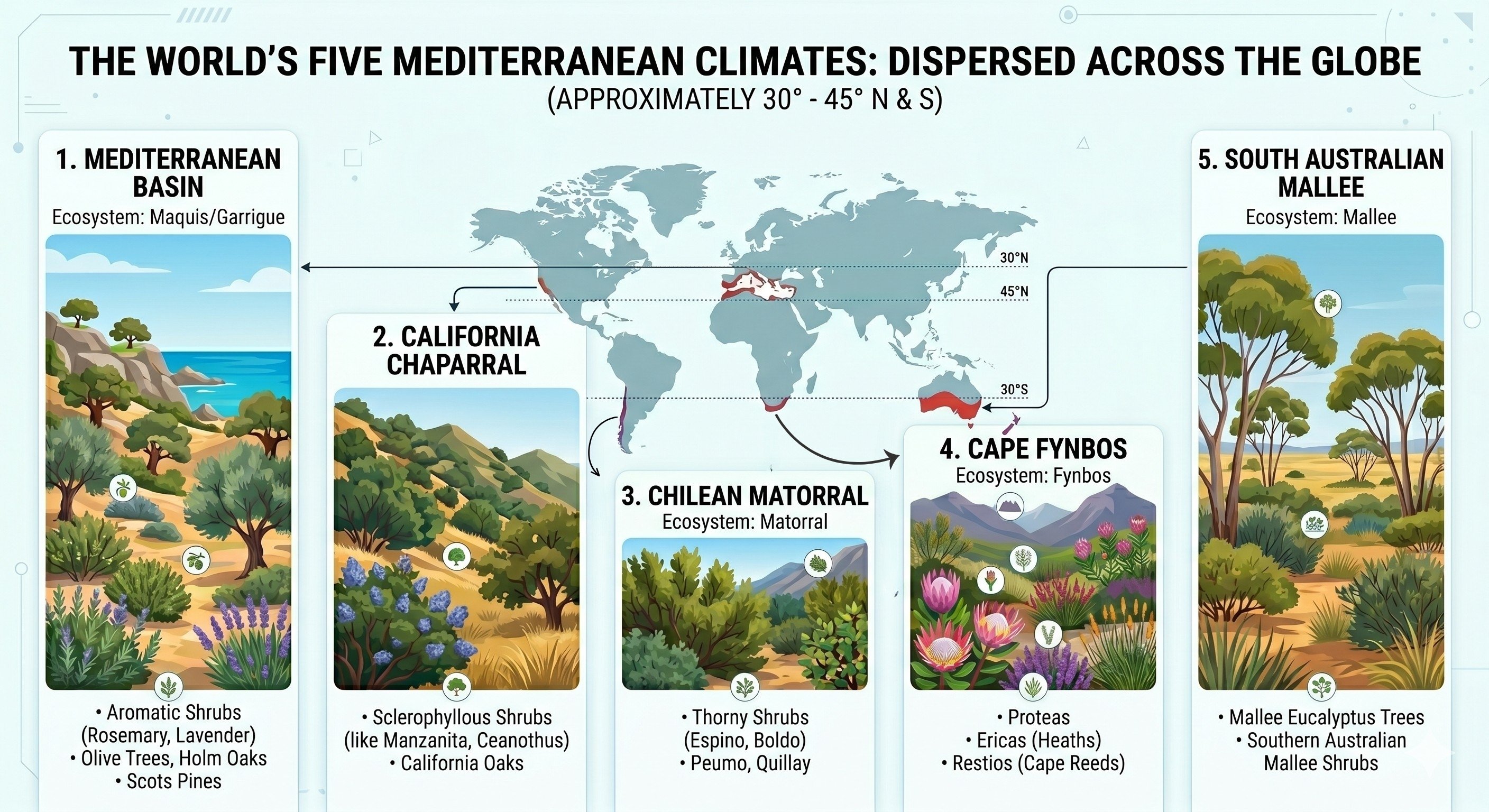
Among the most striking demonstrations that evolution is not merely random is the phenomenon of convergent evolution: the independent arrival, in unrelated lineages, at the same structural or physiological solution to a shared environmental problem. Nowhere is this more legible than in the five Mediterranean-climate zones scattered across the globe — the Mediterranean Basin, the California chaparral, the Chilean matorral, the South African fynbos, and the South Australian mallee. Though separated by thousands of miles and tens of millions of years of independent evolutionary history, each of these regions has produced plant communities that are, in their essential logic, strikingly alike. And yet, in an apparent paradox, the animal communities inhabiting those same landscapes are extraordinarily diverse, differing markedly from one region to the next. Understanding why plants converge while animals diverge turns out to illuminate something fundamental about the relationship between environment, evolution, and the structure of life.
The Problem and Its Parameters
Mediterranean climates are defined by a peculiar inversion. In most of the temperate world, summer brings rain and growth; winter brings dormancy. Here it is the reverse. Moisture arrives in the cool season, when photosynthesis is sluggish and root uptake is slow. The growing season is hot and almost entirely dry. For a plant, this means that the very moment when solar energy is most abundant is the moment when water — the medium through which that energy is metabolically processed — is most scarce. Add to this the periodic sweep of wildfire, which in these landscapes is not a disturbance but a structural feature of the ecosystem, and you have a set of constraints rigorous enough to impose a specific morphology, a specific physiology, and a specific reproductive strategy on any plant that wishes to persist.
The result is what botanists call the sclerophyll: the hard-leaved evergreen. Cork oak (Quercus suber) in the Mediterranean, chamise (Adenostoma fasciculatum) in California, quillay (Quillaja saponaria) in Chile, and the mallee eucalypts of Australia — these are not related plants. But each has arrived at leaves that are small, thick, waxy, and often oriented vertically or curled at their margins, reducing the surface area exposed to the midday sun. Greek sage (Salvia fruticosa) coats its leaves in fine silvery hairs that reflect radiation before it can drive evaporation; the needle-like leaves of rosemary curl downward to trap moisture at their stomatal pores; California’s chamise packs its tiny leaves with flammable resin. These are, functionally, the same invention.
The Underground Wager
Because the above-ground world is so periodically annihilated by fire, the most consequential evolutionary strategy in Mediterranean-climate zones is investment underground. The principle is consistent across all five regions: place the living tissue — the meristems, the nutrient stores, the capacity for regrowth — safely below the soil surface where fire cannot reach it.
In the California chaparral, the manzanita (Arctostaphylos spp.) maintains a massive lignotuber, a dense woody burl from which hundreds of new shoots can erupt within weeks of a fire. The strawberry tree (Arbutus unedo) of the Mediterranean maquis relies on an identical structure — the same answer to the same question, reached through entirely different ancestry. The tree heath (Erica arborea) of the Mediterranean garrigue develops a burl so dense and fire-resistant that it has historically been harvested to make tobacco pipes; its South African relatives, the Cape Heaths (Erica spp., of which the fynbos contains more than six hundred species), do the same thing with the same underground strategy. The South African king protea (Protea cynaroides) resprouts from a thick underground stem after fire; the wild olive (Olea europaea var. sylvestris) regenerates from a gnarled burl if burned to the ground. In Australia, the defining feature of the mallee growth-form is precisely this: multiple trunks arising from a single shared lignotuber that can survive repeated incineration of everything above it.
The geophyte — a plant that survives the hostile season as a dormant bulb or tuber — appears in every one of these zones by the same logic. The mariposa lily (Calochortus spp.) of California, the Chilean alstroemerias, the Cape’s watsonia, the Mediterranean sea squill (Drimia maritima), and the white asphodel (Asphodelus albus) are all unrelated plants practicing the same strategy: retreat entirely underground, wait, and emerge only when fire has passed and nutrients have been liberated into the ash.
Fire as Ally: The Serotinous Seed
Perhaps the most audacious of the convergent strategies is the one that turns fire from an adversary into a reproductive trigger. Serotiny — the retention of seeds in fireproof cones or capsules until heat melts the seal — has evolved independently in each Mediterranean-climate region. The Aleppo pine (Pinus halepensis) holds its seeds in resin-sealed cones that open only when ambient temperatures reach those produced by a wildfire, releasing millions of seeds into a landscape suddenly enriched with ash and stripped of competition. The South African sugarbush (Protea repens) keeps its seeds locked in woody cones for years, waiting for the same signal. The California lilac (Ceanothus spp.) produces seeds that can lie dormant in the soil for decades, germinating only when smoke chemicals penetrate their coats — chemical rather than thermal serotiny, but convergent in function. Chile’s colliguay (Colliguaja odorifera) takes a different mechanical path — its woody capsules explode in summer heat, flinging seeds away from the parent — but the underlying strategy of using fire and heat as a dispersal mechanism is the same. The South Australian mallee eucalypts seal their seeds in woody capsules and release them into the ash.
Alongside these fire-dependent seeders, myrmecochory — seed dispersal by ants — has evolved in parallel in the fynbos and the chaparral. The pincushion protea (Leucospermum cordifolium) coats its seeds in a fatty, nutritious appendage that induces ants to carry them underground, where they are safe from surface fire. California’s ceanothus achieves something similar. The ant becomes, unknowingly, the agent of the plant’s survival strategy.
Aromatic Chemistry as Convergent Physiology
The prevalence of aromatic, resinous, and volatile compounds in Mediterranean-climate plants — rosemary, thyme, mastic, the resins of chamise and spinifex — is itself a case of biochemical convergence. These compounds serve several purposes simultaneously: they deter herbivory (the essential oils of common thyme contain thymol, a powerful antimicrobial and insect repellent), they reduce transpiration by coating leaf surfaces, and they can act as antifreeze in cold winters. That so many unrelated plant lineages — Salvia in the Mediterranean and California, Melaleuca in Australia, Eremophila in the mallee — have converged on volatile aromatic chemistry suggests that this combination of properties is an unusually efficient solution to the Mediterranean climate’s compound demands.
The Exceptions That Prove the Rule
What makes the convergence argument most convincing is that where one region departs from the others, it does so for a reason rooted in its own geological peculiarities. Chile’s matorral is punctuated by columnar cacti (Leucostele chiloensis) and massive terrestrial bromeliads (Puya spp.) that appear nowhere else in the Mediterranean-climate world — but this is because central Chile is adjacent to the Atacama Desert and the Andes, environments that have contributed succulent lineages absent from Europe, South Africa, and Australia. The South African fynbos sits on ancient, extraordinarily nutrient-poor soils; its proteas have evolved specialized root systems — “proteoid roots” — that can extract phosphorus from substrates too impoverished for most plants to exploit. This is a convergence of the second order: a unique local pressure producing a unique local solution, layered atop the broader convergence driven by the Mediterranean climate itself.
The Animal Paradox: Why Diversity Where There Is Convergence?
Against this backdrop of botanical uniformity, the animal life of Mediterranean-climate zones presents an apparent paradox. The lizards, birds, insects, and mammals inhabiting these five regions are not convergent in any comparably systematic way. Each zone has its own characteristic fauna, shaped by its own phylogenetic history. The question of why plants converge so dramatically while animals diversify is, it turns out, as illuminating as the convergence story itself.
The answer begins with a fundamental asymmetry. Plants are sessile. They cannot retreat from a fire, migrate toward water, or behaviorally buffer the Mediterranean summer. Every challenge the climate poses must be solved architecturally — in the structure of the leaf, the chemistry of the bark, the geometry of the root. Animals, by contrast, can move. A lizard can retreat to a crevice during the hottest hours. A bird can shift its territory seasonally. A small mammal can estivate — the summer equivalent of hibernation — in a cool burrow. This behavioral flexibility means that the selective pressure on animal body plans is far less rigid than it is on plant architecture. Animals do not need to reinvent themselves morphologically to survive the Mediterranean summer; they need only to use the landscape intelligently. Behavioral plasticity substitutes for structural convergence, and the result is that animal lineages remain more diverse, more phylogenetically varied, and less constrained to a single morphological template.
The Plant Community as Habitat Mosaic
The extraordinary structural complexity of Mediterranean-climate vegetation — the vertical gradient from phrygana dwarf scrub through garrigue and maquis to open woodland, and the temporal mosaic produced by fire that keeps patches of each stage in continuous rotation — generates an unusually high number of distinct microhabitats within a small area. Each stage of post-fire succession hosts different arthropod communities, different seed stocks, different canopy densities, different amounts of bare ground. A landscape that is never uniformly at one successional stage is, from an animal’s perspective, a landscape that is always offering something. Habitat heterogeneity is one of the most reliable drivers of species richness, and Mediterranean vegetation produces it almost continuously.
Paradoxically, the same fire regime that forces plants into narrow convergent strategies actively promotes animal diversity. A burn immediately creates open ground, abundant ash-fertilized annuals, and a flush of invertebrates exploiting newly available resources — attracting ground-foraging birds and small mammals. The adjacent unburned maquis or mallee simultaneously harbors species dependent on dense cover. Within a few years, the recovering scrub passes through stages of shrub density that favor successively different insectivore and granivore communities. Fire, in other words, does not impose a single state on the landscape but cycles it through many states, each supporting a different animal assemblage. The diversity of animals is partly a diversity of fire-stage specialists.
Insects and the Wealth of Aromatic Chemistry
The resinous, aromatic chemistry that Mediterranean plants evolved to deter generalist herbivores has, over millions of years, become a resource in its own right. Highly specialized insects — particularly beetles, moths, and flies — have evolved the biochemical machinery to detoxify or sequester the very compounds — terpenes, thymol, ladanum — that most animals cannot tolerate. The South African fynbos supports a staggering diversity of specialist pollinators: long-tongued flies, oil-collecting bees, sunbirds, each co-evolved with specific protea or erica flower morphologies. The Cape Floristic Region has been called one of the world’s greatest centers of pollinator diversity precisely because its plant chemistry and flower architecture are so varied and so demanding. What repels a generalist opens a niche for a specialist. The very convergence of plant defensive chemistry, paradoxically, becomes a driver of animal diversification: a landscape saturated with complex secondary compounds is a landscape full of locks, and evolution is endlessly productive of keys.
The Mediterranean Basin’s Ancient Refugial History
In the Mediterranean Basin specifically, there is an additional and quite different explanation for animal diversity: the region served as a glacial refugium throughout the Pleistocene. While northern and central Europe was repeatedly scoured by ice sheets, the Mediterranean’s mild winters allowed populations of birds, reptiles, amphibians, and insects to persist in relative stability. When the ice retreated, those lineages expanded northward — but relict populations remained in the south, accumulating genetic distinctiveness over successive glacial cycles. The result is that the Mediterranean Basin today harbors a disproportionate number of endemic species, particularly in herpetofauna, freshwater fish, and invertebrates — not because the Mediterranean climate is uniquely hospitable in some absolute sense, but because the region has been continuously habitable for long enough to accumulate enormous phylogenetic depth.
The Paradox Resolved
The diversity of animals in Mediterranean-climate zones is produced by a combination of factors that are, in a sense, the inverse of what produces plant convergence. Plants converge because they cannot escape the climate’s demands and must encode solutions in their structure. Animals diversify because they can escape those demands behaviorally, because the structurally complex plant communities plants have built create an exceptionally rich and varied set of niches, and because the aromatic chemical defenses those plants produce create opportunities for specialists willing to crack them. The same selective environment that drives plants toward similarity drives animals toward elaboration.
Convergent evolution does not merely demonstrate that evolution can find the same answer twice. It demonstrates something more profound: that the space of viable biological forms is constrained, that the same selective pressures reliably produce the same morphological and physiological outcomes, and that the history of life is not simply a record of contingent accident but of recurring necessity. The sclerophyllous leaf, the underground burl, the serotinous cone, the aromatic resin — these are not arbitrary inventions but discoveries, made independently by lineages separated by oceans and epochs, of the only ways to live well in a world of summer drought and periodic fire. And yet it is precisely because plants have made those discoveries so thoroughly — have built landscapes of such structural and chemical complexity — that animals have been liberated to proliferate into the niches those plants create. Convergence and diversification turn out to be two sides of the same ecological coin: the plant’s constraint becomes the animal’s opportunity.